Česká zemědělská univerzita
Katedra genetiky a obecné zootechniky
Autor:
Kateřina Hamanová
Název:
Polymorfismus DNA se zaměřením na mikrosatelity u koní
(zkrácený obsah disertační práce – podklad pro seminář určený pro
chovatelskou veřejnost)
Cíle:
Na základě vysoce polymorfních mikrosatelitních markerů, doporučených
pro standardní protokol testování paternity u koní podle mezinárodních
srovnávacích testů ISAG, 1997(98), byla charakterizována a porovnávána různá
plemena koní se zaměřením na genové rezervy ČR.
Vybraný panel markerů byl ověřován u jednotlivých plemen z hlediska:
· jeho vypovídací schopnosti
· testování paternity pomocí sekvenátoru v našich podmínkách
· schopnosti stanovení odpovídajících hodnot heterozygotnosti a genetické distance.
Charakterizace současného stavu jednotlivých populací
(plemen) na základě zjištěných heterozygotností a genetických distancí.
Co to je mikrosatelitní marker ?
·
Mikrosatelitní DNA neboli
mikrosatelity, také nazývané STR
(short tandem repeats – krátká tandemová
opakování), jsou krátké segmenty DNA, ve kterých se mnohokrát opakují
specifické motivy nukleotidových sekvencí.
·
Základem těchto repetitivních
úseků jsou sekvence o velikosti od 1 do
4 až 6 bp.
·
Motiv je obvykle
opakován 60 krát a velikost jednotky
se pohybuje zhruba v rozpětí 20 – 100bp.
·
Typický sledovaný lokus má více než 10 alel.
·
Heterozygotnost těchto
lokusů je obvykle vyšší než 0.60, a
to dokonce i v relativně malých
skupinách zvířat.
·
Díky jejich extrémní
variabilitě (polymorfismu) a relativně snadné detekci, jsou považovány za jedny
z nejvhodnějších genetických markerů.
·
Navíc k jejich vysoké
variabilitě, jsou mikrosatelity také velmi
hojně rozšířené v eukaryotickém genomu, což je staví do pozice
preferovaných markerů genetického mapování s vysokou citlivostí rozlišení.
·
Ačkoliv se mikrosatelity
vyskytují v genomu ve velkém množství, jejich biologická funkce je stále nejasná.
·
V koňském genomu je
jeden z nejhojněji zastoupených repetičních motivů (TG)n motiv, který vykazuje značně velký stupeň polymorfismu a jeho frekvence je odhadována zhruba 1 na 100 000 bp v genomu koní. Oproti tomu
frekvence (TC)n repetic je vůči
zmíněným (TG)n repeticím daleko
nižší, i když jsou zhruba stejně polymorfní.
·
Rozlišit určité mikrosatelitní jednotky v tomto
množství nelze přirozeně, ani na základě charakteru motivu (ten je uniformní),
ani na základě počtu repetic motivu (určitý počet se může v genomu
opakovat třeba stokrát). Základní charakteristikou mikrosatelitů jsou proto
jedinečné sekvence nukleotidů, přiléhající nebo vymezující mikrosatelitní
sekvenci tzv. „flanking sequences„.
Tyto přiléhající sekvence určují jedinečnou pozici mikrosatelitů v genomu.
·
Každý jedinec má dvě kopie
každého mikrosatelitu, jeden zděděný po matce, druhý po otci. To, že je délka
každého mikrosatelitu děděna mendelisticky, dělá mikrosatelity ideálními
polymorfními markery při mapování genomu, testování paternity a identifikaci.
·
Mikrosatelity velmi rychle
nahrazují metody spočívající na RFLP a RAPD
(random amplified polymorphic DNA = náhodně
amplifikovaná polymorfní DNA) ve většině aplikací v populační
biologii, od identifikování příbuzných jedinců, až po odvozování demografických
parametrů.
·
Pomocí unikátních primerů je
amplifikována, tedy zmnožena určitá oblast genomu, jež zahrnuje definovanou strukturu
opakování, která je zodpovědná za sledovanou odchylku.
·
Mikrosatelitní mapy jsou nyní
dostupné téměř u všech organismů genetického a ekonomického zájmu, od člověka,
myši, skotu, prasete, ovci, slepici, drozofilu, až po rajče, sóju, rýži a mnoho
dalších.
·
V kontrastu
s jejich důležitostí ve vnitrodruhových studiích, mikrosatelity dosud
nevytvořily žádný skutečný příspěvek pro fylogenetickou rekonstrukci.
C. Metodika
Bylo použito těchto 12 markerů (viz. tab), doporučených konferencí ISAG.
Lokus
|
Chromozom |
Sekvence
primeru
|
|
VHL20 |
30
|
5’-CAA GTC CTC
TTA CTT GAA GAC TAG-3’ 5’-AAC TCA GGG
AGA ATC TTC CTC AG-3’ |
|
HTG4 |
9 |
5’-CTA TCT CAG
TCT TGA TTG CAG GAC-3’ 5’-CTC CCT CCC
TCC CTC TGT TCT C -3’ |
|
HTG6 |
15 |
5’-CCT GCT TGG AGG
CTG TGA TAA GAT-3’ 5’-GTT CAC TGA
ATG TCA AAT TCT GCT-3’ |
|
HTG7 |
4 |
5’-CCT GAA GCA
GAA CAT CCC TCC TTG-3’ 5’-ATA AAG TGT
CTG GGC AGA GCT GCT -3’ |
|
HTG10 |
21 |
5’-CAA TTC CCG
CCC CAC CCC CGG CA-3’ 5’-TTT TTA TTC
TGA TCT GTC ACA TTT -3’ |
|
AHT4 |
24 |
5’-AAC CGC CTG
AGC AAG GAA GT-3’ 5’-GCT CCC AGA
GAG TTT ACC CT-3’ |
|
AHT5 |
8 |
5’-ACG GAC ACA
TCC CTG CCT GC-3’ 5’-GCA GGC TAA
GGG GGC TCA GC-3’ |
|
ASB2 |
15 |
5’-CCA CTA AGT
GTC GTT TCA GAA GG-3’ 5’-CAC AAC TGA
GTT CTC TGA TAG G-3’ |
|
HMS2 |
|
5’-CTT GCA GTC
GAA TGT GTA TTA AAT G-3’ 5’-ACG GTG GCA
ACT GCC AAG GAA G-3’ |
|
HMS3 |
9 |
5’-CCA ACT CCT
TGT CAC ATA ACA AGA-3’ 5’-CCA TCC TCA
CTT TTT CAC TTT GTT-3’ |
|
HMS6 |
4 |
5’-GAA GCT GCC
AGT ATT CAA CCA TTG-3’ 5’-CTC CAT CTT
GTG AAG TGT AAC TCA-3’ |
|
HMS7 |
1 |
5’-CAG GAA ACT
CAT GTT GAT ACC ATC -3’ 5’-TGT TGT TGA
AAC ATA CCT TGA CTG T -3’ |
Celkem bylo testováno 5 plemen koní (267 zvířat), která byla dále porovnávána s dalšími plemeny koní, uvedenými v literatuře, pomocí níže uvedených statistických ukazatelů, na základě jejich alelických frekvencí. Všechna tato plemena vzájemně vykazují více či méně rozdílů v tělesné stavbě, temperamentu, možnostech využití, … a původu.
Testovaná
zvířata:
·
SN
… slezský norik - 39 ks, farma Klokočov na Moravě
(spol.
PROMED OPAVA s.r.o.)
·
LN
… lánský norik - 16 ks, školní statek Lány
·
A1/1
… anglický plnokrevník - 133 ks (různé)
·
CT
… český teplokrevník - 18 ks (různé)
·
Z
… zemaituka - 17 ks (Litva)
·
H
… hucul - 44 ks, - farma Hucul –
Janova Hora (33)
-
Hucul klub – Laka u Kokořína (11)
Dále na základě alelických frekvencí
porovnáváno se zvířaty:
·
QH
… Quarter horse „čtvrtkař“ - 8850 ks (Bowling et al.,1997)
·
(Cothran et al., 1998)
![]() KB … kladrubský bělouš - 87 ks
KB … kladrubský bělouš - 87 ks
·
KV
… kladrubský vraník - 74 ks
·
![]() ECAtb … Equus Caballus – Thoroughbred =
ECAtb … Equus Caballus – Thoroughbred =
(Breen et
al., 1994)
domestikovaný kůň -
plnokrevník - 90 ks …
·
ECAmix
… kříženci domestikovaného koně - 40 ks … ..
·
EPR
… kůň Převalského - 50 ks … … … … … … … …
Pomocí polymerázové řetězové reakce (PCR) prováděné na
automatickém termálním cykleru, bylo v multiplexních i jednoduchých
polymerázových řetězových reakcích namnoženy (amplifikovány) dané DNA úseky za
použití uvedených primerů .
Polymerázová řetězová reakce
zahrnuje:
n horký start 10min. při teplotě 95°C
n 30 cyklů, kde byly prováděny všechny kroky po 1min.:
- denaturace při teplotě 93°C
- anelace při teplotě 55°C
- extenze při teplotě 72°C
n prodloužená extenze (anelace)
a) 10min. při teolotě 72°C – pro lokusy množené a typizované jednotlivě
b) 60 min. při teolotě 72°C – pro lokusy množené a typizované v multiplexu
n chlazení 4°C
DNA typizace jedinců
Zastoupení jednotlivých genotypů mikrosatelitů bylo
zjišťováno pomocí automatického sekvenátoru ABI-PRISM 310 Perkin-Elmer, program
GeneScan Analysys a princip typizace spočívá v:
1. Produktů
PCR všech amplifikovaných lokusů se umístí do DNA sekvenátoru (tedy
automatizovaného elektroforetického detektoru jedné linie). Vzorky DNA molekul
díky elektrickému poli migrují od katody k anodě, skrze laserový detektor.
GENESCAN software, běžící na PC značky Mackintosh, shromažďuje signály, a poté
stanoví velikost (bp) pro každý vzorek. Data z GENESCANu jsou přímo
exportována do software Genotyper pro automatickou genotypizaci. Pomocí více
barevného fluorescenčního značení je umožněno vzorky hustě naskládat do
multiplexu bez ohledu na to, zda-li se vzorky svými alelickými velikostmi
překrývají. Pomocí interního standardu,
který běží spolu se vzorky v každém běhu je pak docilována
maximální přesnost při určování velikostí jednotlivých alel. Veškerá data jsou
elektronicky uchovávána.
Tato DNA
typizace byla prováděna v imunologické laboratoři ČMSCH s.r.o.
v Hradišťku pod Medníkem.
|
Počet alel na lokus |
|||||||
Lokus
|
A1/1 |
SN |
LN |
Z |
CT |
H |
Ř |
|
VHL20 |
8 |
8 |
7 |
8 |
5 |
9 |
7,5 |
|
HTG4 |
5 |
4 |
4 |
6 |
3 |
6 |
4,7 |
|
AHT4 |
4 |
6 |
4 |
5 |
5 |
10 |
5,7 |
|
HMS7 |
5 |
6 |
5 |
4 |
7 |
7 |
5,7 |
|
HTG6 |
5 |
3 |
4 |
5 |
7 |
5 |
4,8 |
|
AHT5 |
6 |
7 |
6 |
3 |
5 |
7 |
5,7 |
|
HMS6 |
4 |
3 |
6 |
5 |
4 |
6 |
4,7 |
|
ASB2 |
13 |
6 |
5 |
5 |
8 |
7 |
7,3 |
|
HTG10 |
8 |
6 |
6 |
5 |
7 |
8 |
6,7 |
|
HTG7 |
6 |
4 |
4 |
4 |
4 |
5 |
4,5 |
|
HMS3 |
7 |
6 |
6 |
6 |
8 |
7 |
6,7 |
|
HMS2 |
6 |
6 |
7 |
5 |
6 |
5 |
5,8 |
Plemeno hucul vykazuje ve srovnání s ostatními testovanými plemeny vyšší hodnoty co do počtu zjištěných alel.
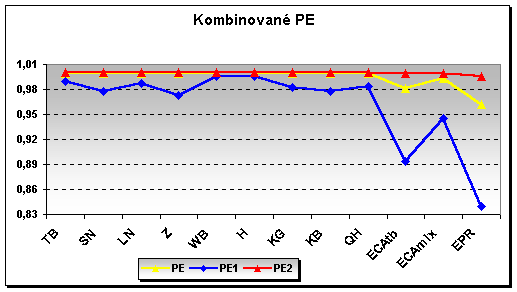
Z hlediska testování paternity (PE) vykazují uvedené markery jednu z nejvyšších pravděpodobností právě u plemene hucul, bez ohledu na to zda byli testováni oba rodiče, či pouze jeden z nich. (viz graf) PE – standardní ověřování na základě obou rodičů, obdobně jako novodobější metoda PE2, PE1 – testování paternity za nepřítomnosti jednoho z rodičů.
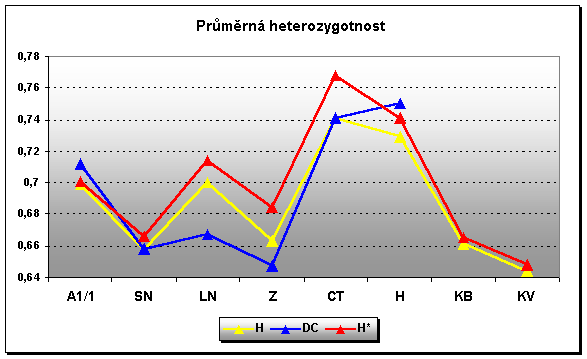
Průměrná heterozygotnost patří mezi ukazatele charakterizující stav heterozygotů v populaci. Jednoduše čím vyšší je uvedené číslo tím lépe. Hodně zjednodušeně řečeno, existují jisté hladiny tohoto parametru při jejichž naplnění můžeme považovat populaci z hlediska inbrední deprese ve stavu nedotčeném příbuzenskou plemenitbou a na naopak. Takovými hladinami jsou pro hodnoty průměrné He u mikrosatelitů 0,8 a 0,5. Z toho vyplývá pro plemeno hucul velmi dobrý výsledek na rozdíl třeba od kladrubského vraníka, kde hodnoty jednoznačně potvrdily skutečnost značného vlivu příbuzenské plemenitby, apod. Samozřejmě to nemůžeme brát za dogma, protože v takovém případě by bylo třeba sledovat daleko větší počet markerů.
¨Tři varianty heterozygotností H – standardní heteroz. na základě alelických frekvencí, DC – heteroz. vycházející přímo z heterozygotních jedinců a H* - nevychýlená heteroz. zameřená na skupiny s malým počtem testovaných zvířat.
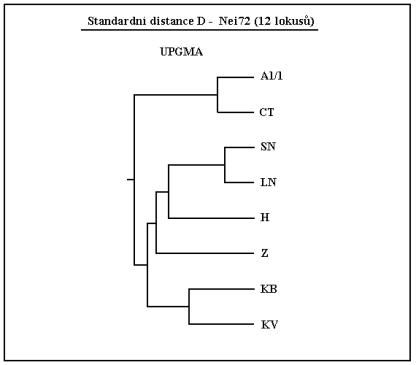
Ukazatelem porovnávajícím plemena jsou genetické distance. Celá řada takovýchto parametrů existuje, z nichž každý vyhovuje trošku jiným podmínkám. V našem případě se ukázala jako nejméně vychýlená distance od NEIE 1972.
A spolu s UPGMA metodou shlukování pak vytváří následující dendrogram. Pro fylogenetické stromy není 12 markerů příliš mnoho, nicméně vzhledem k ceně stanovení musí pro orientaci postačit.
|
Populace |
A1/1 |
SN |
LN |
Z |
CT |
H |
KB |
KV |
|
|
|
|
|
|
|
|
|
|
|
A1/1 |
***** |
0,020 |
0,003 |
0,026 |
0,017 |
0,017 |
0,021 |
0,033 |
SN |
0,547 |
***** |
0,014 |
0,014 |
0,012 |
0,019 |
0,033 |
0,030 |
|
LN |
0,486 |
0,122 |
***** |
0,021 |
0,006 |
0,016 |
0,032 |
0,024 |
|
Z |
0,694 |
0,405 |
0,377 |
***** |
0,021 |
0,016 |
0,038 |
0,030 |
|
CT |
0,153 |
0,407 |
0,371 |
0,546 |
***** |
0,014 |
0,018 |
0,035 |
|
H |
0,447 |
0,352 |
0,293 |
0,479 |
0,414 |
***** |
0,018 |
0,021 |
|
KB |
0,568 |
0,409 |
0,484 |
0,488 |
0,411 |
0,564 |
***** |
0,021 |
|
KV |
0,672 |
0,371 |
0,512 |
0,434 |
0,621 |
0,594 |
0,273 |
***** |
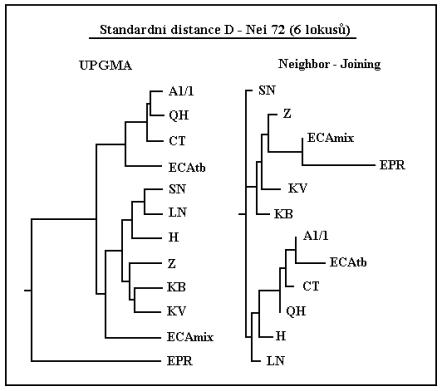
Tento složitější dendrogram pak vychází z méně početného panelu markerů, zato jsou v něm zahrnuta i další plemena. Obdobně jako na předešlém i zde je postavení hucula nevýrazné a pro přesně jší určení by bylo skutečně potřeba daleko více markerů.
Zveřejněno s laskavým souhlasem autorky.